|
|
| (не показано 7 промежуточных версий 4 участников) |
| Строка 1: |
Строка 1: |
| − | {{DISPLAYTITLE:Адаптация мышц к физической нагрузке}} | + | {{SportFiz}} |
| − | {{Эндокринология}}
| + | == Адаптация как биологическая основа эффектов занятий физическими упражнениями == |
| − | == Механизмы адаптации скелетных мышц к физическим упражнениям == | |
| | | | |
| − | [[Гипертрофия мышц]]
| + | '''Адаптация''' — это способность организма приспосабливаться к внешней среде или изменениям в самом организме. Различают процесс и состояние адаптации. Состояние адаптации — это состоявшаяся физиологическая адаптация. Оно характеризуется устойчивым уровнем активности и взаимосвязи систем органов, тканей и механизмов регуляции, обеспечивающих нормальный уровень жизнедеятельности организма в новых условиях внешней и внутренней среды. Это состояние достигается в течение определенного времени, за которое происходит адаптация (процесс адаптации) (Ключевые факторы адаптации..., 1986; Медведев, 1984; Солодков, Сологуб, 2003). |
| | | | |
| − | Физические упражнения можно рассматривать в качестве комплексного физиологического стимула, который воздействует на различные аспекты клеточной функции. Скелетные мышцы представляют одну из тканей, которая реагирует на физические упражнения, подвергаясь ряду видоизменений на уровне некоторых своих компонентов. Скорость сокращения скелетной мышцы, количество создаваемой при сокращении силы, а также способность противостоять утомлению, — все это важные показатели, имеющие непосредственное отношение к спортивным показателям. Благодаря высокой лабильности различных характеристик мышечной ткани, таких, как размер фибрилл, состав фибрилл и степень капилляризации ткани, скелетные мышцы способны должным образом приспосабливаться к изменениям, возникающим в ходе тренировочного процесса. В то же время характер адаптации скелетных мышц к силовым упражнениям и упражнениям на выносливость будет отличаться, что свидетельствует о существовании различных систем реагирования на нагрузку. Таким образом, приспособительный процесс скелетных мышц к тренировочным нагрузкам можно рассматривать как совокупность согласованных локальных и периферических событий, ключевыми регуляторными сигналами к которым являются гормональные, механические, метаболические и нервные факторы. Изменения в скорости синтеза гормонов и ростовых факторов, а также содержание их рецепторов являются важными факторами регуляции приспособительного процесса, позволяющего скелетным мышцам удовлетворить физиологические потребности различных видов двигательной активности. Краткое описание роли некоторых гормонов и ростовых факторов в гипертрофии мышц, в регуляции фенотипа миофибрилл и преобразовании капиллярной кровеносной сети и составляет содержание данной главы.
| + | '''По механизмам возникновения''' различают такие виды адаптации: |
| | | | |
| − | === Увеличение скелетной мышцы: роль ядер мышечной клетки и миосателлитоцитов ===
| + | *генотипическая, обусловленная врожденными механизмами функционирования систем и возникшая на основе наследственности, изменчивости, естественного отбора, мутаций и являющаяся характерной для вида живых организмов; |
| | | | |
| − | Усиление синтеза сократительных белков является безоговорочным условием увеличения размера мышечных клеток в ответ на тренировочную нагрузку. В процессе [[Рост мышц|роста скелетных мышц]] изменяется не только интенсивность синтеза белка, но и скорость его деградации (Goldbeig et al., 1975). У человека усиление синтеза белка выше уровня покоя происходит очень быстро, в течение 1 — 4 ч после завершения разового тренировочного занятия (Wong, Booth, 1990; Chcsley ct al., 1992; Biolo ct al., 1995; Philips ct al., 1997). В начале мышечной гипертрофии усиление синтеза белка коррелирует с ростом активности РНК (Laurent et al., 1978; Wong, Booth, 1990). Передача мРНК облегчается теми факторами, активность которых, как известно, регулируется путем их фосфорилирования (Frederickson, Sonebcig, 1993; Wada ct al., 1996). Параллельно с этими изменениями после тренировочного занятия происходит усиление транспорта аминокислот в мышцы, подвергавшиеся нагрузке. С теоретической точки зрения это увеличивает доступность аминокислот для белкового синтеза (Biolo et al., 1997).
| + | *фенотипическая, приобретаемая в течение индивидуальной жизни и проявляемая относительно влияния определенного фактора внешней среды (температура, АД, влажность, физические нагрузки и др.). |
| | | | |
| − | Ряд данных свидетельствует о том, что после этого первоначального этапа необходимым условием продолжения гипертрофии мышц является увеличение уровня РНК (в отличие от увеличения активности РНК, происходившего вначале). Здесь возросшее количество мРНК может быть обусловлено либо усилением генной транскрипции в клеточных ядрах, либо увеличением количества ядер. Мышечные волокна взрослого человека содержат сотни ядер и каждое ядро осуществляет синтез белка в каком-то ограниченном объеме цитоплазмы, получившем название "ядерный компонент” (Cheek, 1985; Hall, Ralston, 1989; Allen ct al., 1999). Важно отметить, что хотя ядра мышечной клетки прошли митоз, они способны обеспечивать увеличение фибрилл лишь до определенного предела, после которого становится необходимым привлечение новых ядер. Это предположение подтверждается результатами исследований человека и животных, демонстрирующими, что гипертрофия скелетных мышечных волокон сопровождается значительным увеличением количества ядер (Goldberg et al., 1975; Cabric, James, 1983; Winchester, Gonyea, 1992; Allen et al., 1995; Kadi, 2000). У хорошо тренированных людей, например у тяжеловесов, количество ядер в гипертрофированной фибрилле скелетной мышцы больше, чем у лиц, ведущих малоподвижный образ жизни. Установлено существование линейной зависимости между количеством ядер и площадью поперечного сечения миофибриллы (Kadi et al., 1999а; Kadi, 2000). Появление новых ядер в увеличившейся миофибрилле играет роль в поддержании постоянного ядерно-цитоплазматического соотношения, т. е. стабильного размера ядерного компонента. О появлении новых ядер в гипертрофирующихся миофибриллах сообщалось для лиц разного возраста (Hikida et al., 1998; Kadi, Tomcll, 2000).
| + | '''По срокам возникновения''' различают срочную (быструю, незавершенную) и долговременную (медленную, завершенную) фазы адаптации. |
| | | | |
| − | В то время как при гипертрофии мышц наблюдается увеличение количества ядер в мышечной клетке, при исследованиях процессов атрофии на животных было отмечено противоположное явление. Уменьшение количества ядер происходит в результате атрофии мышечных волокон после перерезки спинного мозга (Allen et al., 1995), космических полетов (Allen ct al., 1996) или временного обездвиживания задней конечности (Hikida et al., 1997). Таким образом, изменение количества ядер на мышечную клетку, по-видимому, имеет большое значение для регуляции размера клеточной фибриллы. В то же время необходимо иметь в виду, что увеличение количества ядер в мышечном волокне будет происходить до тех пор, пока активность превращения уже существующих ядер окажется способной для обеспечения роста их количества. Действительно, заметные изменения количества ядер в мышечном волокне наблюдались в мышцах, гипертрофированных более чем на 26 % (Cabric, James, 1983; Allen et al., 1995; Roy ct al., 1999; Kadi, Torncll, 2000), но не в гипертрофированных на 6,8—15,5 % (Giddings, Gonyea, 1992).
| + | '''Срочная адаптация''' — это реакция организма на действующий раздражитель. Ее реализация происходит на основе сформировавшихся ранее физиологических механизмов. Адаптация реализуется «с места» и не является совершенной. Примером срочной адаптации являются физиологические реакции организма на выполнение физической нагрузки: повышение ЧСС, ЧД, АД и других показателей. Работа требует затрат энергии, и для ее реализации усиливается деятельность вегетативных систем. |
| | | | |
| − | Поскольку ядра дифференцированного мышечного волокна неспособны делиться, основным источником новых ядер в гипертрофированных мышечных волокнах являются миосателлитоциты или клетки-спутники (Moss, Lcblond, 1971; Schiaffino et al., 1976). Миосателлитоциты расположены между базальной пластиной и плазматической мембраной мышечных подокон (Mauro, 1961), для них характерно высокое ядерно-цитоплазматическое соотношение, хорошо развитый аппарат Гольджи, выраженный гранулярный эндоплазматический ретикулум и гетерохроматинизированное ядро (Campion, 1984). Активация клеток-спутников может происходить под воздействием ряда стимулов, после чего они начинают активно делиться. После этого образовавшиеся в результате митоза дочерние клетки сливаются с расположенными рядом дифференцированными мышечными клетками, обеспечивая таким образом увеличение количества ядер. Роль образующихся при делении миосателлитоцитов ядер в процессе мышечной гипертрофии подтверждается также экспериментами на животных моделях, демонстрирующими, что для обеспечения роста мышц необходимы активация и последующая пролиферация клеток-спутников (Rosenblatt, Parry, 1992; Rosenblatt et al., 1994).
| + | '''Долговременная адаптация''' возникает в результате многократного влияния фактора, то есть многократной реализации срочной адаптации. Вследствие постепенного количественного накопления изменений организм приобретает новое качество — превращается в адаптированный, и потому одни и те же движения выполняются экономнее, эффективнее, а максимальная работа выводит организм на более высокий уровень функционирования. |
| | | | |
| − | Показано, что параллельно с гипертрофией мышц интенсивная силовая тренировка индуцирует существенное увеличение количества клеток-спутников в скелетных мышцах (Kadi, 2000; Roth et al., 2001). Сообщалось об увеличении на 46 % доли миосателлитоцитов в скелетной мышце молодой женщины после 10 недель силовой тренировки (Kadi, Tomell, 2000). Не так давно увеличение количества миосателлитоцитов было обнаружено в скелетных мышцах группы мужчин в возрасте 70—80 лет, занимавшихся тренировкой выносливости (Charifi et al., 2003а). Таким образом, клетки-спутники обеспечивают увеличение количества ядер в мышечном волокне и возобновление своего собственного пула (Bischoff, 1994; Schultz, McCormick, 1994; Kennedy et al., 1988; Yamada et al., 1989; McCormick, Schultz, 1992; Antonio, Gonyea, 1993; Kadi, Thomell, 1999). Вновь сформированные мышечные волокна замещают поврежденные или вносят свой вклад в гиперплазию мышечных волокон, только если количество вновь сформированных волокон превышает количество волокон, поврежденных во время тренировки.
| + | Выполнение оптимальных, адекватных индивидуальным возможностям организма, физических нагрузок обеспечивает развитие структурных изменений прогрессивного характера — '''рациональной адаптации''', что способствует развитию резервных возможностей организма спортсменов. Чрезмерные физические нагрузки, превышающие функциональные возможности организма, вызывают нарушение функциональных связей между его структурными уровнями и ведут к развитию '''нерациональной адаптации'''. В отличие от рациональной, нерациональная адаптация характеризуется стремительным становлением и может сопровождаться дистрофическими изменениями тканей, а впоследствии и структурно-функциональными нарушениями органов и тканей (Алексаняйц, 2003; Козлов, 1997; Спортивная медицина..., 2003). |
| | | | |
| − | === Влияние андрогенных анаболических стероидов ===
| + | '''Срочная адаптация''' рассматривается как состояние общего напряжения организма, возникающее вследствие влияния очень сильного раздражителя. Термин «[[стресс]]» впервые использовал канадский ученый Ганс Селье в 1936 г. Он показал, что во время влияния на организм стрессового раздражителя возникает активизация деятельности организма, увеличивающая секрецию АКТГ, которая стимулирует прежде всего деятельность коры надпочечников. |
| | | | |
| − | Результаты исследований, проведенных на животных, показали, что использование андрогенных [[Анаболические стероиды|анаболических стероидов]] сопровождается значительным увеличением размера мышц и мышечной силы (Egginton, 1987; Salmons, 1992). Применение [[Тестостерон|тестостерона]] в концентрациях, превышающих физиологические, у мужчин с различным уровнем физической подготовленности на протяжении 10 недель сопровождалось существенным увеличением мышечной силы и поперечного сечения четырехглавой мышцы бедра (Basin et al., 1996). Известно, что андрогенные [[Анаболические гормоны|анаболические стероиды]] увеличивают интенсивность синтеза белка и способствуют росту мышц как in vivo, так и in vitro (Powers, Florini, 1975; Rogozkin, 1979). У человека прием анаболических стероидов на протяжении длительного времени усиливает степень гипертрофии мышечных волокон у хорошотренированных тяжелоатлетов (Kadi et al., 1999b). Скелетные мышцы тяжелоатлетов, принимавших анаболические стероиды, характеризуются экстремально большим размером мышечных волокон и большим количеством ядер в мышечных клетках (Kadi et al., 1999b). Подобную картину наблюдали на животных моделях, в частности, было обнаружено, что андрогенные анаболические стероиды опосредуют свое миотрофное воздействие путем увеличения количества ядер в мышечных волокнах и увеличения количества мышечных волокон (Galavazi, Szirmai, 1971; Sassoon, Kelley, 1986; Joubcrt, Tobin, 1989; Joubert, Tobin, 1995). Таким образом, анаболические стероиды способствуют увеличению количества ядер с целью обеспечения белкового синтеза в чрезвычайно гипертрофированных мышечных волокнах (Kadi et al., 1999b). Основным механизмом, посредством которого андрогенные анаболические стероиды индуцируют мышечную гипертрофию, является активация и индукция пролиферации миосателлитоцитов, которые впоследствии сливаются с уже существующими мышечными волокнами или между собой, формируя новые мышечные волокна. С таким выводом согласуются результаты иммуногистохимической локализации рецепторов андрогенов в культивируемых клетках-спутниках, демонстрирующие возможность непосредственного воздействия анаболических стероидов на миосателлитоциты (Doumit et al., 1996).
| + | [[Гормоны]] коры [[Надпочечники|надпочечников]] стимулируют приспособительные механизмы, благодаря которым организм адаптируется к влиянию раздражителя. Механизмы такой срочной адаптации являются общими для разных стрессовых влияний — физических, химических, эмоциональных и др. В результате возникло понятие — общий адаптационный синдром (Дубровский, 2005; Фомин, Вавилов, 1991). |
| | | | |
| − | == [[Андрогенные рецепторы]] ==
| + | '''Общий адаптационный синдром''' — это комплекс неспецифических реакций организма на действие раздражителя, происходящих в несколько стадий: тревоги, резистентности, истощения. |
| | | | |
| − | Блокада рецепторов [[Андроген|андрогенов]] оксендолоном — антагонистом рецепторов андрогенов — подавляет гипертрофию, стимулированную физическими упражнениями (Inoue et al., 1994). Хотя в процессах адаптации мышечных волокон к физическим упражнениям принимает участие несколько различных факторов, результаты этого исследования однозначно показывают, что рецепторы андрогенов являются важными посредниками в индуцированной физическими нагрузками мышечной гипертрофии.
| + | '''Стадия тревоги''' характеризуется предельной мобилизацией функций. Физиологические механизмы этой стадии характерны для срочной адаптации, реализующейся по типу стресс-реакции с максимальной мобилизацией вегетативных функций. |
| | | | |
| − | Рецепторы андрогенов принадлежат к семейству гормончувствительных регуляторов превращения. В случае связывания гормонов с рецептором он переходит в активированное состояние и комплекс гормон-рецептор перемещается к гормончувствительному элементу в ядре. Связывание с определенными генами увеличивает уровень их транскрипции (Luke, Coffey, 1994). В ранних работах сообщалось, что андрогенные рецепторы локализуются в цитозоле мышечной клетки (Krieg, 1976; Max et al., 1981). Использование имммуногистохимии со специфическими поли- и моноклональными антителами позволило установить локализацию рецепторов андрогенов практически во всех типах тканей (Sar et al., 1990; Takedaetal., 1990; Ruizeveld-De-Winter et al., 1991; Kimura et al., 1993; Janssen et al., 1994). В скелетной мышце рецепторы андрогенов содержатся в ядрах мышечных волокон (Takeda et al., 1990; Kimura et al., 1993; Dorlochter et al., 1994; Kadi et al., 2000b) и в миосателлитоцитах (Doumit et al., 1996). В норме в покоящихся мышечных волокнах человека расположение рецепторов андрогенов происходит лишь в некоторых, а не во всех ядрах (Kadi et al., 2000b). Сообщалось также о различиях в содержании рецепторов андрогенов па мышечное волокно между трапециевидной и латеральной широкой мышцами бедра (Kadi, 2000). Похожие различия во внутримышечном содержании рецепторов андрогенов были также продемонстрированы для мышц лягушки (Dorlochter et al., 1994). С данными о различиях внутримышечного содержания рецепторов андрогенов согласуются результаты исследований, где было показано, что наибольшая чувствительность скелетных мышц морской свинки к стимуляции тестостероном наблюдается в области головы и шеи, а при перемещении к задней части туловища происходит ее постепенное снижение (Kochakian, Tillotson, 1957). Различия во внутримышечном содержании рецепторов андрогенов могут отражать различия в эмбриологическом происхождении, иннервации и функциональных потребностях различных мышц.
| + | '''Стадия резистентности''' характеризуется активным поиском устойчивого состояния. Ее механизмы являются основой долговременной адаптации. |
| | | | |
| − | Физические тренировки оказывают влияние на количество мест связывания андрогенов в скелетных мышцах. Увеличение количества рецепторов андрогенов должно приводить к возрастанию чувствительности мышц к андрогенам, циркулирующим в системе кровообращения. Исследования на животных показали, что существенное увеличение количества рецепторов андрогенов происходит в результате тренировки мышечной силы и выносливости, а также электростимуляции мышц (Inoue et al., 1993; Deschenes et al., 1994). Амплитуда изменений количества рецепторов андрогенов, обусловленного физической тренировкой, варьирует в различных мышцах (Hickson, Kurowsky, 1986; Deschenes ct al., 1994; Kadi et al., 2000b). Занятия силовыми упражнениями в течение продолжительного времени влекут за собой изменения количества ядер мышечных волокон, содержащих рецепторы андрогенов, в трапециевидной мышце, но не в латеральной широкой мышце бедра человека (Kadi et al., 2000b). Аналогичные различия в регуляции количества рецепторов андрогенов после занятий физическими упражнениями существуют между длинным разгибателем пальцев ноги, камбаловидной, икроножной и подошвенной мышцами крысы (Hickson, Kurowsky, 1986; Salmons, 1992; Inoue etal., 1993; Bricout et al., 1994; Deschenes et al., 1994).
| + | '''Стадия истощения''' проявляется в случае, когда сила действующего раздражителя продолжает расти, превышая функциональные и метаболические возможности организма, возникает срыв адаптации, переходящий в дезадаптацию. При оптимальной организации тренировочного процесса стадия резистентности может не возникать в течение продолжительного времени. |
| | | | |
| − | Было проведено изучение влияния силовой и аэробной тренировки на содержание рецепторов андрогенов и их сродство к дигидротестостерону в медленных и быстрых мышечных волокнах у крысы (Deschenes et al., 1994). Ни один из видов физической тренировки не вызывал изменений чувствительности рецепторов андрогенов к дигидротестостерону. [[Аэробные упражнения|Аэробная тренировка]] индуцировала увеличение способности связывать андрогены в медленных мышечных волокнах, тогда как силовая тренировка сопровождалась аналогичными изменениями в быстрых волокнах. Таким образом, характер изменений содержания рецепторов андрогенов зависит не только от типа мышцы, но и от вида двигательной активности (тренировка выносливости или силовые упражнения). Изменения количества рецепторов андрогенов в ответ на физическую тренировку происходят достаточно быстро. После трех дней электростимуляции икроножной мышцы у крыс наблюдается увеличение количества рецепторов андрогенов в стимулированной мышце на 25 %, которое сопровождается прогрессивной гипертрофией мышцы (Inoue et al., 1993).
| + | == Адаптация к физической нагрузке == |
| | + | {{Теория спортивной тренировки}} |
| | + | [[Image:21vek3.jpg|250px|thumb|right|Действие закона адаптации в процессе тренировки спортсменов (по Zatsiorsky, 1995)]] |
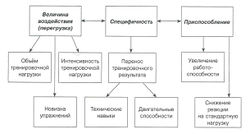
| | + | Целенаправленный тренировочный процесс приводит к разнообразным изменениям в организме спортсмена и, таким образом, увеличивает их [[Спортивная_работоспособность|работоспособность]]. С биологической точки зрения [[Спортивная тренировка|тренировка]] - это длительный процесс адаптации спортсмена к различным нагрузкам. Следовательно, упражнения, тренировки и различные задания служат стимулами для адаптации. В биологии адаптация рассматривается как процесс приспособления, который происходит в организме под воздействием изменяющихся условий жизни. Вообще говоря, адаптация, первоначально описанная великим физиологом Гансом Селье (1950), является одним из фундаментальных законов науки о жизни. Профессор В.М. Зациорский применил принципы процесса адаптации к спортивной тренировке (1995); он обнаружил, что приспособление спортсменов к увеличивающимся нагрузкам обусловлено тремя главными факторами: величиной воздействия, его спецификой и приспособляемостью спортсмена (рис.). |
| | | | |
| − | Проведена также оценка влияния андрогенных анаболических стероидов на содержание рецепторов андрогенов. На животных моделях в культуре миосателлитоцитов было показано, что андрогенные анаболические стероиды могут повышать (Doumit et al., 1996) либо понижать (Lin et al., 1993; Bricout et al., 1994) количество рецепторов андрогенов в мышечной клетке. Установлено, что прием тестостерона усиливает иммунореактивность рецептора андрогенов в ядрах миосателлитоцитов свиньи (Doumit et al., 1996). Однако с помощью радиоактивного конкурентного анализа было показано, что после применения андрогенных анаболических стероидов происходит снижение концентрации рецепторов андрогенов в камбаловидной мышце и длинном разгибателе пальцев ноги (Bricout et al., 1994). В действительности, влияние андрогенных анаболических стероидов на содержание рецепторов андрогенов также может зависеть от типа клеток. Так, волокна скелетных мышц лягушки из плечевой области характеризуются наибольшей чувствительностью к тестостерону по сравнению с волокнами из других участков тела (Regnier, Herrera, 1993а, 1993b). Скелетные мышцы кролика, которые характеризуются сходным размером мышечных волокон, соотношением различных их типов, иннервацией и кровоснабжением могут сильно отличаться характером своего ответа на воздействие андрогенными анаболическими стероидами (Salmons, 1992). И наконец, у человека самостоятельный прием андрогенных анаболических стероидов на протяжении длительного времени вызывал изменения в ядрах, содержащих рецепторы андрогенов, мышечных волокон трапециевидной мышцы, при этом в латеральной широкой мышце бедра подобных изменений не наблюдалось (Kadi et al., 2000b). Очевидно, что для лучшего понимания изменения количества рецепторов андрогенов в скелетных мышцах в ответ на физиологические и нефизиологические воздействия необходимо проведение дальнейших исследований.
| + | По закону адаптации эффективная тренировка должна обеспечивать оптимальную комбинацию этих трёх главных факторов, а она, в свою очередь, определяет прогресс в работе над [[Физическая работоспособность|работоспособностью]] спортсменов. Обобщая сказанное, можно назвать упомянутые выше факторы принципами адаптации применительно к тренировочному процессу. |
| | | | |
| − | == [[Факторы роста фибробластов]] == | + | === Величина тренировочной нагрузки и принцип перегрузки === |
| | | | |
| − | Семейство факторов роста фибробластов (ФРФ) включает 10 представителей, участвующих в реализации различных биологических функций (Yamaguchi, Rossant, 1995). Некоторые изоформы ФРФ играют важную роль в процессе увеличения мышечных волокон в ответ на физиологические стимулы, другие могут оказывать свое миотрофное воздействие в ответ на физиологические стимулы, миотрофная роль третьих может проявляться во время восстановления мышечных волокон после повреждения мышцы. В этом отношении наиболее важным фактором, необходимым для нормальной регенерации скелетной мышцы после повреждения, является ФРФ6 (Floss et al., 1997), который может принимать участие в важных регенеративных событиях, таких, как активация и пролиферация клеток-спутников и содержание важных миогенных регуляторных факторов. На модели птиц, у которых утяжеляли крыло, было показано увеличение активности ФРФ2 и ФРФ4, локализованных в дифференцированной скелетной мышце в периферическом матриксе миофибрилы, которое сопровождало вызванную растягиваниями гипертрофию (Mitchell et al., 1999). Стимуляция ФРФ2 и ФРФ4 наряду со специфической локализацией подтверждает их роль в активации и пролиферации клеток-спутников (Mitchell et al., 1999). На основании расположения в том же месте, что и клетки-спутники, было выдвинуто предположение о том, что высвобождение этих факторов от гепариновых компонентов может иметь определенное значение в формировании новых мышечных волокон после физической тренировки (Yamada et al., 1989). Изменения в характере взаимодействия ФРФ и гепаринсульфатпротеогликанов в мышцах, подвергающихся физической нагрузке, может модулировать доступность ФРФ для миосателлитоцитов.
| + | [[Тренировочная нагрузка]] вызывает реакцию спортсмена и служит стимулом для адаптации. Величина воздействия может регулироваться тремя факторами: объёмом нагрузки, её интенсивностью и новизной упражнений. Важно отметить, что рост уровня подготовленности может быть достигнут, только если величина воздействия достаточна. '''Принцип перегрузки''' гласит, что для увеличения уровня подготовленности требуется применение нагрузки (воздействия), величина которой превышает привычный уровень. |
| | | | |
| − | Исследование механизмов, управляющих превращением механических нагрузок в ростовые процессы скелетных мышц, позволило установить, что высвобождение ФРФ2 усиливается параллельно с увеличением мышечной нагрузки в модели культуры дифференцированных клеток скелетной мышцы человека (Clarke, Feeback, 1996). При нейтрализации биологической активности ФРФ2 происходило ингибирование ростовых процессов. Этот эксперимент однозначно подтверждает, что высвобождение ФРФ2 является важным аутокринным механизмом влияния механической нагрузки как фактор стимуляции ростовых процессов скелетной мышцы (Clarke, Feeback, 1996).
| + | В соответствии с принципом перегрузки величина нагрузки имеет первостепенное значение и должна тщательно оцениваться и программироваться. Общий подход к описанию величины нагрузки представлен ниже (табл.). |
| | | | |
| − | == [[Инсулиноподобные факторы роста и их рецепторы]] ==
| + | '''Характеристики величины нагрузки''' |
| | | | |
| − | Выработка различных изоформ [[Инсулиноподобный фактор роста|инсулиноподобных факторов роста]] (ИФР) происходит во многих тканях и имеет важное значение для развития организма в эмбриональном и постнаталыгам периодах: ИФР-II необходим для нормального развития плода, в то время как [[ИФР-1|ИФР-I]] имеет важное значение для пре- и постнатального роста (DeChiara et al., 1990; Baker et al., 1993). Скелетные мышцы, выполняющие работу, вырабатывают и используют ИФР-1 (Brahm et al., 1997), который считается важным фактором, опосредующим увеличение скелетных мышц в ответ на физическую тренировку. ИФР-1 способен стимулировать пролиферацию, дифференцировку и слияние миосателлитоцитов (Dodson et al., 1985; Florini et al., 1991; Quinn et al., 1994; Goldspink D.F. et al., 1995). Иммуногистохимические исследования показали, что расположение ИФР-1 происходит в клетках-спутниках и в мышечных трубочках регенерирующих скелетных мышц крысы (Jennische et а)., 1987; Jennische, 1989; Jennische, Matejka, 1992).
| + | <table border="1" style="border-collapse:collapse;" cellpadding="3"> |
| | + | <tr><td bgcolor="e5e5e5"> |
| | + | <p>Компонент тренировочной нагрузки</p></td><td bgcolor="e5e5e5"> |
| | + | <p>Основные показатели</p></td><td bgcolor="e5e5e5"> |
| | + | <p>Возможные индикаторы</p></td></tr> |
| | + | <tr><td> |
| | + | <p>Объём</p></td><td> |
| | + | <p>Сумма всех выполненных упражнений, представленная количественной характеристикой</p></td><td> |
| | + | <p>Общее количество тренировок за период времени, например, за неделю, месяц, год и т.д.</p> |
| | + | <p>Общее время, затраченное на тренировки за данный период.</p> |
| | + | <p>Общий километраж за тренировочный период.</p> |
| | + | <p>Общее количество подъёмов, бросков, прыжков и пр. за тренировочный период</p></td></tr> |
| | + | <tr><td> |
| | + | <p>Интенсивность</p></td><td> |
| | + | <p>1) Интенсивность рабочей нагрузки.</p> |
| | + | <p>2) Сумма упражнений, выполненных с увеличенной мощностью</p></td><td> |
| | + | <p>Уровень мощности (%) относительно максимума.</p> |
| | + | <p>Уровень мощности, определяемый по величине ЧСС.</p> |
| | + | <p>Соответствие определённой зоне интенсивности. Частный объём упражнений, выполненных с увеличенной мощностью (километраж, затраченное время, количество попыток и т.д.)</p></td></tr> |
| | + | <tr><td> |
| | + | <p>Новизна</p> |
| | + | <p>упражнения</p></td><td> |
| | + | <p>Наличие упражнения, которое содержит неизвестные элементы или детали / новые комбинации известных элементов</p></td><td> |
| | + | <p>Количество новых (или относительно новых) упражнений, включённых в программу тренировки</p></td></tr> |
| | + | </table> |
| | | | |
| − | После интенсивного выполнения эксцентрических упражнений в передней большеберцовой мышце крысы происходит увеличение иммунореактивности ИФР-1, которое наблюдается главным образом в мышечных волокнах (Yan et al., 1993). У человека интенсивные физические тренировки индуцируют увеличение иммунореактивности ИФР-1 в широкой медиальной мышце бедра (Hellsten et al., 1996).
| + | '''Объём тренировочной нагрузки'''. Исторически самый простой способ увеличить нагрузку состоял в увеличении объёма тренировок. У высококвалифицированных спортсменов во многих видах спорта в 1930-х годах количество тренировок в неделю равнялось 2-3, в 1960-х увеличилось до 6-8, а в 1980-х достигло 9-14. С тех пор частота тренировок осталась на том же уровне. В течение долгого времени считалось, что желание увеличить объём тренировок было ограничено физиологическими и социальными факторами. С точки зрения физиологов уже был достигнут верхний предел резервов человека; социологи же высказывали беспокойство о том, что кроме тренировок спортсмены нуждаются в образовании, профессии, личной жизни и т.д. |
| | | | |
| − | Иммуногистохимическая окраска локализует ИФР-1 в ядрах мышечных клеток или клеток-спутников (Hellsten et al., 1996). Повышение уровня мРНК и белка ИФР-1, происходящее параллельно с гипертрофией мышечных волокон, показывает, что этот ростовой фактор, влияющий на миосателлитоциты, занимает ключевое место в процессе увеличения мышечных волокон (Adams, Haddad, 1996). Показано, что у человека уровень мРНК ИФР-1 повышается через 48 ч после одноразового комплекса концентрических и эксцентрических физических нагрузок медиальной широкой мышцы бедра (при этом увеличение мРНК после эксцентрической нагрузки более важно) (Bamman et al., 2001). В этой же работе было обнаружено существенное увеличение количества мРНК рецепторов андрогенов, происходящее после концентрической или эксцентрической нагрузки.
| + | Несмотря на это, объём тренировочных нагрузок в мировом спорте стремился к увеличению до конца 1980-х годов. Этот объём стабилизировался и даже уменьшился только в течение двух прошедших десятилетий. В любом случае, увеличение тренировочной нагрузки - слишком очевидный фактор личного прогресса спортсмена в любом виде спорта. Оценка объёма тренировочной нагрузки - обычная практика в видах спорта на выносливость, где выполненный километраж традиционно подсчитывается, однако это может стать трудной задачей в игровых видах или в единоборствах, где не просто суммировать количество специфических спортивных действий. |
| | | | |
| − | Развитие молекулярной биологии позволило открыть новые изоформы ИФР-1 (Yang et al., 1996; Goldspink G., 1999; Hameed et al., 2002). Описаны два основных типа ИФР-1, которые располагаются в скелетных мышцах. Первая изоформа имеет системный характер действия, обладает сходством с основным вырабатываемым в печени ИФР-1 и носит название мышечный ИФР-1 печеночного типа (Yang et al., 1996). Вторая мышечная изоформа, получившая название механозависимый фактор роста (МФР), была обнаружена в скелетных мышцах, подвергавшихся растягиванию и чрезмерным нагрузкам, и обладает аутокринным и паракринным действием (Yang et al., 1996). В ответ на растягивания путем иммобилизации нижней конечности в разогнутом положении наблюдали существенное увеличение МФР в длинном разгибателе пальцев стопы кролика (Yang et al., 1996). Позднее было обнаружено, что после растягивания и растягивания в сочетании с электростимуляцией с частотой 10 Гц в длинном разгибателе пальцев стопы кролика наблюдается также существенное повышение уровней мРНК ИФР-1 печеночного типа и МФР, однако после одной электростимуляции подобных изменений не происходило (МсКоу et al., 1999). Эти эксперименты ясно показывают, что изоформы ИФР-1 опосредуют увеличение скелетной мышцы преимущественно в ответ на возросшую механическую нагрузку. Роль МФР в гипертрофии мышц исследовали у лиц зрелого и старшего возраста. После 10 этапов из 6 повторений упражнения для сгибателя колена, которое выполнялось одной ногой с нагрузкой 80 % 1 повторного максимума (1ПМ), существенное увеличение мРНК МФР наблюдали только у лиц зрелого возраста (Hameed et al., 2003). Был сделан вывод о том, что ослабление изменений уровня МФР в ответ па интенсивные занятия силовыми упражнениями у лиц старшего возраста может свидетельствовать о возрастном сниженим чувствительности мышечных волокон к механической нагрузке.
| + | '''Интенсивность тренировочной нагрузки'''. Интенсивность тренировочной нагрузки обычно рассматривается в двух аспектах: |
| | | | |
| − | Применение ИФР-1 было предложено для лечения различных нейромышечных заболеваний, характеризующихся атрофией мышц. В частности, ИФР-1 успешно использовали для предотвращения развития индуцированной приемом стероидов мышечной атрофии (Kanda et al., 1999). На клеточном уровне было показано, что ежедневный прием [[Соматотропин|соматотропиого гормона]] и ИФР-1 в сочетании с чрезмерной функциональной нагрузкой камбаловидной мышцы у крысы приводит к увеличению мышцы одновременно с увеличением количества ядер в мышечных волокнах (McCall et al., 1998).
| + | *как мерило уровня мощности относительно максимума (иногда относительно уровня соревновательной мощности); |
| | + | *как составляющая общего объёма тренировочной нагрузки, который выполнен с увеличенной (выше обычного) мощностью. |
| | | | |
| − | Стимулирующее ростовые процессы воздействие ИФР-1 и ИФР-П осуществляется при участии рецептора ИФР-1. Мыши с нарушениями функции рецептора ИФР-1 погибали вскоре после рождения (Baker et al., 1993). Отсутствие рецептора ИФР-1 вызывает у мышей тяжелую гипоплазию, что свидетельствует о необходимости рецептора ИФР-1 для формирования дифференцированных скелетных мышц (Baker et al., 1993). В то время как функциональная инактивация рецептора ИФР-1 вызывала выраженную гипоплазию мышц, а также снижение уровней MyoD и миогенина (два важных представителя миогенных регуляторных факторов) (Fernandez et al., 2002), при сверхэкспрессии рецепторов наблюдаются противоположные явления (Quinn et al., 1994). Таким образом, рецептор ИФР-1 в данный момент рассматривается в качестве главного регулятора мышечной массы, регулирующего активность специфических для мышечной ткани генов. Однократное интенсивное занятие физическими упражнениями сопровождается существенным увеличением связывающей способности и чувствительности рецептора ИФР-1, а также повышением уровня мРНК рецептора в скелетных мышцах крысы (Willis et al., 1997). Точно так продолжительные физические тренировки вызывают заметное увеличение концентрации рецепторов ИФР-1 и [[инсулин]]а (Willis et al., 1998). В целом эти данные однозначно подтверждают важную роль ИФР-1 и рецепторов ИФР-1 в гипертрофии мышц.
| + | Конечно, более интенсивные упражнения вызывают более явную реакцию в организме спортсмена. Следовательно, интенсивность нагрузки оценивается как показателями внешней нагрузки ([[Скорость (двигательное физическое качество)|скоростью]], [[Мощность мышц|мощностью]], поднятыми весами), так и посредством индикаторов реакции организма. [[Частота сердечных сокращений (ЧСС)]], например, является одним из широко распространённых показателей физиологической реакции. ЧСС обеспечивает достаточную индикацию уровня интенсивности широкого спектра упражнений. |
| | | | |
| − | == [[Кровоснабжение скелетных мышц]] ==
| + | В последние годы '''зоны интенсивности''' (ЗИ) стали широко использоваться во многих видах спорта и для [[Планирование спортивной тренировки|планирования]], и для посттренировочной оценки (Viru, 1995). В соответствии с этим подходом весь диапазон интенсивности подразделяется на зоны (обычно их пять). Каждая ЗИ описывается рядом значимых индикаторов, каждый из которых отражает диапазон показателей, соответствующих этой зоне. Обычно для характеристики определённой зоны интенсивности используются лактат крови, ЧСС, скорость (или время работы, или мощность) и темп движений. За прошедшее десятилетие в связи с развитием новых спортивных технологий (таких как мониторы ЧСС, портативные анализаторы лактата крови, электронные измерительные системы времени) этот подход был существенно усовершенствован. |
| | | | |
| − | Кровеносные сосуды скелетных мышц формируют обширную сеть капилляров вокруг мышечных волокон. Капилляры состоят из одинарного слоя эндотелиальных клеток, которые со стороны просвета сосуда покрыты гликокаликсом (обогащенный углеводами периферический участок клеточной мембраны), а с обратной — несут базальную мембрану. Капиллярная сеть, которая представляет собой своеобразный “конец” сердечно-сосудистой системы, играет важную роль в обеспечении мышечных волокон питательными веществами и осуществлении обмена кислорода и диоксида углерода в мышцах. Показано, что при тренировке выносливости в различных скелетных мышцах человека происходит изменение расположения кровеносных сосудов (Andersen Р., Henriksson, 1977; Hudiicka et al., 1992; Wang et al., 1993; Kadi et al., 2000a; Charifi et al., 2003b).
| + | '''Новизна упражнения'''. Новизна упражнения является третьим компонентом, определяющим величину тренировочной нагрузки; реакция спортсменов весьма зависима от того, насколько привычными являются для них некоторые упражнения. Однако в отличие от объёма и интенсивности новизна упражнения редко рассматривается как фактор, влияющий на тренировочную нагрузку. Известно, что творчески настроенные тренеры повсюду ищут новые оригинальные упражнения, чтобы обогатить существующий набор и сделать тренировочный процесс более привлекательным. Эффект применения этих новшеств проявляется в более выраженной физиологической реакции спортсмена. |
| | | | |
| − | === Фактор роста сосудистого эндотелия ===
| + | '''Пример'''. ''Игорь Кошкин (СССР), один из всемирно известных экспертов в области плавания, который тренировал трёхкратного олимпийского чемпиона Владимира Сальникова, сказал другим тренерам: «Если Вы начнёте использовать стояние на голове как упражнение для ваших пловцов, начальный эффект будет существенным и положительным из-за его новизны. Но этот эффект будет очень кратковременным, потому что это упражнение не затрагивает специфические плавательные способности ваших спортсменов».'' |
| | | | |
| − | Фактор роста сосудистого эндотелия (vascular endothelial growth factor, VEGF) представляет собой гепаринсвязывающий специфический для эндотелиальных клеток митоген, который стимулирует ангиогенез в различных тканях. Как физические нагрузки, так и гипоксия могут вызывать увеличение уровня мРНК VEGF в скелетной мышце человека (Gustafsson et al. 1999; Richardson et al., 1999). Повышение уровня мРНК VEGF у нормальных здоровых людей и страдающих заболеваниями ночек происходит уже после одного занятия физическими упражнениями с нагрузкой, равной 50 % максимальной нагрузки (Wagner et al., 2001). Кроме того, при гипоксии и физической нагрузке наблюдается увеличение количества рецепторов VEGF (flt-1 и flk-1) (Tagaki et al., 1996; Gerber et al., 1997; Olfert et al., 2001). Таким образом, фактор роста сосудистого эндотелия VEGF и его рецепторы участвуют в увеличении плотности капиллярной сети в ответ на двигательную активность. У нетренированных людей уже после одного занятия физическими упражнениями наблюдается значительный рост уровня мРНК VEGF в скелетных мышцах (Richardson et al., 2000). Эти события могут отражать первоначальный интенсивный рост капилляров в нетренированных мышцах, происходящий в начале выполнения тренировочной программы, после чего процессы ангиогенеза замедляются и требуют для своей стимуляции увеличения тренировочной нагрузки, поскольку уровень тренированности мышцы повышается.
| + | Это замечание подчёркивает сложность проблемы, связанной с новизной упражнения. Действительно, нетрудно найти упражнение, с которым спортсмены не знакомы, но нелегко найти незнакомое им упражнение, которое соответствует специфическим по виду спорта физиологическим, биомеханическим и психологическим требованиям. Именно поэтому специфичность тренировочной нагрузки, которая будет рассмотрена ниже, является обязательным фактором адаптации в спортивной тренировке. |
| | | | |
| − | В число факторов, контролирующих экспрессию VEGF, входит составляющая индуцируемого гипоксией фактора 1 (hypoxia inducible factor 1, HIF-1), который в настоящий момент рассматривается в качестве основного фактора регуляции. Влияние кратковременной тренировки на VEGF и HIF-1 изучали у 8 здоровых мужчин. Несмотря на увеличение уровня VEGF после 7 тренировочных занятий, не было обнаружено никаких изменений уровня мРНК составляющей HIF-1 (Gustafsson et al., 2002). Это свидетельствует о том, что увеличение количества мРНК HIF-1, возможно, не единственный фактор регуляции содержания VEGF при занятиях физическими упражнениями. Вместе с тем воздействие различных факторов, контролирующих рост капиллярной сети, вероятнее всего, происходит в различные моменты времени, что существенно затрудняет оценку их вклада, если только исследователь не проводит наблюдения за этим адаптивным процессом в целом.
| + | === Специфичность тренировочной нагрузки === |
| | | | |
| − | Предполагается, что факторы роста фибробластов (ФРФ) также играют роль в аигиогенезе в скелетных мышцах. Однако последние исследования показали, что их вклад в процесс роста капилляров гораздо менее значительный по сравнению с VEGF (Richardson et al., 2000; Wagner et al., 2001). В настоящий момент, хотя и считается, что VEGF — наиболее важный фактор ангиогенеза, принимающий участие в адаптации капиллярной сети в скелетных мышцах человека, необходимы дальнейшие исследования, направленные па расширение наших знаний о вкладе всех известных факторов ангиогенеза в формирование и рост кровеносной сети скелетной мышцы.
| + | Как видно из рисунка, специфичность тренировочной нагрузки характеризуется [[Перенос тренированности|переносом результата тренировки]] с одного задания (выполнения вспомогательного упражнения) на другое (основное упражнение). Обычно тренеры используют широкий набор упражнений, большинство из которых может быть разделено на две группы: |
| | | | |
| − | == [[Типы мышечных волокон]] ==
| + | *упражнения для совершенствования физических качеств (силы, выносливости и др.); |
| | + | *упражнения для совершенствования технических навыков. |
| | | | |
| − | Существование различных типов мышечных волокон обеспечивает значительную гетерогенность тканей скелетных мышц и их способность выполнять разнообразные функциональные задачи. Иммуногистохимический и биохимический анализ скелетных мышц показал, что такое структурно-функциональное разнообразие мышечных волокон обусловлено существованием широкого спектра изоформ миозина. Миозин — молекула, от которой наряду с актином зависит мышечное сокращение. Молекула миозина состоит из двух тяжелых цепей (МуНС) и четырех легких цепей (MyLC) (Schiaffino, Reggiani, 1996; Pette, Staron, 1997). Тяжелые цепи миозина представлены несколькими изоформами, от свойств которых зависят скоростно-силовые качества мышечных волокон.
| + | Конечно, эти упражнения могут быть скомбинированы, чтобы улучшить взаимодействие между физическими качествами и техническими навыками. В любом случае, полезность каждого упражнения зависит от того, как оно влияет на выполнение главного (соревновательного) упражнения. Другими словами, перенос двигательных способностей и перенос технических навыков с тренировочного упражнения на соревновательное определяет, насколько полезными являются эти вспомогательные упражнения. |
| | | | |
| − | В скелетных мышцах взрослого человека происходит экспрессия четырех наиболее важных изоформ МуНС: MyHCip, MyHCIIA, MyHCIIX/IID и МуНСПВ. Каждая изоформа характеризуется специфической скоростью сокращения и развиваемым усилием. Волокна, содержащие MyHCI, отличаются низкой скоростью сокращения и развивают меньшее усилие по сравнению с волокнами, содержащими MyHCIIA, ИХ и IIB. Среди волокон, состоящих их быстрых МуНС, наиболее быстрыми и сильными являются те, которые построены из МуНСПВ, за ними следуют волокна, в состав которых входят МуНСИХ и MyHCIIA (Bottineli et al., 1994a, 1994b).
| + | === Аккомодация === |
| | | | |
| − | Занятия физическими упражнениями могут приводить к существенным изменениям сократительных свойств скелетных мышц. Принято считать, что тренировка выносливости сопровождается увеличением количества медленных изоформ миозина (Baumann et al., 1987; Schaub et al., 1989). В то же время силовая тренировка вызывает увеличение MyHCIIA и уменьшение МуНСПХ (Staron et al., 1991; Adams et al., 1993; Andersen J.L. et al., 1994; Fry et al., 1994; Kraemer et al., 1995; Kadi, Thorncll, 1999; Andersen J.L., Aagaard, 2000). Кроме того, предполагается, что мышечные волокна, содержащие МуНСИХ, у основной массы людей очень редко вовлекаются в выполнение работы в процессе обычной ежедневной активности. Если они начинают вовлекаться в выполнение работы, например в процессе физической тренировки, то превращаются в волокна, содержащие MyHCIIA (волокна, включающие эту изоформу тяжелых цепей миозина, обладают большей выносливостью по сравнению с волокнами типа ИХ) (Goldspink G. et al., 1991; Staron et al., 1991; Kraemer et al., 1995). Во время тренировки мышечной силы или выносливости происходит значительное изменение гормонального фона скелетных мышц, которое является мощным сигналом, способным запустить процесс изменения содержания изоформ миозина в мышцах, подвергающихся физической нагрузке.
| + | Две тесно связанные особенности характеризуют аккомодацию (обязательный компонент процесса адаптации, вызванного тренировкой): |
| | | | |
| − | === Влияние тестостерона ===
| + | *увеличение работоспособности; |
| | + | *уменьшение реакции на стандартную физическую нагрузку. |
| | + | [[Image:21vek4.jpg|250px|thumb|right|Изменение скорости анаэробного порога (сплошная линия) и накопление лактата крови после прохождения 1000-метровой дистанции с заранее определённой скоростью (пунктирная линия) у квалифицированных байдарочников в ходе подготовительного периода]] |
| | + | Увеличение работоспособности может быть отслежено с помощью специфических по виду спорта показателей, таких как результаты выполнения работы до отказа, скорость анаэробного порога в видах спорта на выносливость и т.д. Стандартная физическая нагрузка может быть организована при обследовании спортсменов на эргометре или при тестировании их с заранее заданной скоростью или мощностью. Оба варианта можно проследить на примере результатов наблюдения за подготовкой байдарочников мирового класса в течение одного сезона. |
| | | | |
| − | В некоторых экспериментах на животных после применения андрогенных анаболических стероидов наблюдали изменение соотношения изоформ тяжелых цепей миозина в сторону увеличения медленных изоформ (Fritzshe et al., 1994; Czesla ct al., 1997). Сообщалось об увеличении доли волокон, содержащих MyHCIIA, наряду с сокращением количества волокон, содержащих МуНСПВ, в ряде скелетных мышц грызунов после применения андрогенных анаболических стероидов (Eggington, 1987; Dimauro et al., 1992). Однако сообщалось также о том, что андрогенные стероиды вызывают уменьшение доли мышечных волокон, содержащих MyHCIIA, по отношению к волокнам, состоящим из МуНСПВ (Kelly et al., 1985; Lyons et al., 1986; Salmons, 1992). Эти результаты говорят о том, что характер воздействия андрогенных анаболических стероидов на сократительные способности может зависеть от типа мышц и у различных видов может быть разным. Действительно, существуют и другие данные, свидетельствующие об отсутствии какого-либо воздействия андрогенных анаболических стероидов па соотношение мышечных волокон, содержащих различные изоформы МуНС. Например, в экспериментах на животных чрезмерная нагрузка мышц вызывала увеличение содержания медленных MyHCI, и дополнительное использование андрогенных анаболических стероидов не влияло на характер содержания тяжелых цепей миозина (Boissonneault et al., 1987). Точно так прием андрогенных анаболических стероидон не вызывал изменений сдвига соотношения изоформ МуНС, вызванного экспериментами с обездвиживанием нижней конечности (Tsika et al., 1987). Наконец, не удалось обнаружить никаких различий в соотношении разных изоформ МуНС в трапециевидной мышце хорошо тренированных тяжелоатлетов, принимавших и не принимавших андрогенные анаболические стероиды (Kadi et al., 1999b).
| + | '''Конкретный пример'''. ''Группа из девяти квалифицированных байдарочников обследовалась в течение одного подготовительного сезона. Выполнялся тест на воде (4 раза по 1000 м) с возрастающей нагрузкой для определения скорости анаэробного порога. Кроме того, выполнялась стандартная нагрузка - гребля на дистанции 1000 м на заранее определённой скорости, соответствующей средней интенсивности (скорость была запрограммирована лидирующей моторной лодкой). После испытания брались пробы крови. Графики показывают значительное увеличение скорости анаэробного порога у всех спортсменов и аналогичное уменьшение накопления лактата крови, вызванное греблей с постоянной скоростью (рис.).'' |
| | | | |
| − | === Влияние эстрогенов ===
| + | Этот пример показывает, что процесс аккомодации у спортсменов может контролироваться посредством испытаний, выполняемых как с максимальными, так и со стандартными усилиями. Этот подход может также использоваться в видах спорта, где спортивный результат не поддаётся измерению, таких как игры с мячом, где стандартная нагрузка может быть запрограммирована определённой комбинацией специфических по виду спорта элементов с фиксированной частотой выполнения и диапазоном движения. |
| | | | |
| − | Хорошо известен тот факт, что уменьшение развиваемой силы происходит в менопаузе (Greeves et al., 1999; Dionne et al., 2000; Meeuwsen et al., 2000). Ha клеточном уровне показано, что удаление яичников сопровождается изменением соотношения изоформ тяжелых цепей миозина в сторону увеличения медленных волокон и понижением спонтанного бега у крыс (Kadi et al., 2000). В целом изменения соотношения изоформ МуНС имеют следующую тенденцию: МуНС I < IIA < ИХ < ИВ. Эти результаты можно интерпретировать как то, что после удаления яичников происходят общие изменения содержания изоформ МуНС в сторону преобладания медленных изоформ, специфическая стимуляция генов медленных изоформ МуНС, или специфическое подавления генов, кодирующих медленные изоформы МуНС. Если животным с удаленными яичниками позволяли бегать и вводили эстрогены, в составе МуНС не происходило никаких изменений (Kadi et al., 2000). Таким образом, можно предположить, что двигательная активность в сочетании с введением эстрогенов способствуют поддержанию исходного соотношения быстрых и медленных волокон в мышцах.
| + | У процесса аккомодации есть много субъективных индикаторов: с увеличением работоспособности спортсмены сообщают о большей «свободе движения», облегчении дыхания во время длительной работы, лучшем расслаблении мышц, усилении специфических по виду спорта ощущений, подобных «чувству воды» в водных видах, «чувству льда» в катании на коньках и т.д. Все эти субъективные оценки очень важны и для тренера, и для спортсмена; желательно отмечать их в дневниках спортсменов и журналах тренеров. |
| | | | |
| − | === Влияние соматотропного гормона ===
| + | В заключение надо отметить, что общая логика принципов адаптации может быть представлена в такой последовательности: |
| | | | |
| − | Сообщается о том, что прием соматотропного гормона (СТГ) индуцирует увеличение количества МуНСИХ в латеральной широкой мышце бедра у здоровых мужчин старшего возраста (Lange et al., 2002). Изменение соотношения изоформ МуНС в сторону увеличеиия МуНСИХ авторы исследования рассматривали как “омоложение” состава тяжелых цепей миозина, поскольку старение обычно сопровождается уменьшением доли МуНСИХ в этой группе мышц (Lange et al., 2002). Однако доля МуНСИХ у пациентов с дефицитом СТГ была выше по сравнению с основной массой здорового населения (Daugaard et al., 1999). Более того, после лечения больных с дефицитом СТГ препаратами рекомбинантного гормона роста в течение 6 месяцев у них не было выявлено никаких изменений в соотношении различных ияоформ МуНС (Daugaard et al., 1999). Аналогичным образом было показано, что применение СТГ у крыс приводит к существенному увеличению поперечного сечения мышечных волокон типа II в камбаловидной мышце, не оказывая заметного влияния на содержание различных изоформ в составе мышечных волокон (Aroniadou-Anderjaska et al., 1996). Вопрос о том, приводит ли повышение уровня СТГ к изменению соотношения изоформ МуНС в сторону увеличения быстрых изоформ миозина, требует дальнейших исследований.
| + | *тренировка с адекватной рабочей нагрузкой вызывает желаемые реакции в организме спортсменов (принцип величины воздействия), |
| | + | *эти реакции вызывают процесс приспособления, который приводит к увеличению работоспособности и более экономному реагированию на стандартные рабочие нагрузки (принцип аккомодации); |
| | + | *увеличенный уровень работоспособности отражается на выполнении соревновательного упражнения в соответствии с тренировочными результатами, перенесёнными с различных упражнений на главное соревновательное (принцип специфичности). |
| | | | |
| − | === Влияние гормонов щитовидной железы === | + | Нарушая эти взаимосвязи, мы понижаем [[Тренировочные эффекты|тренировочный эффект]], и чем выше уровень спортсмена, тем значительнее будет ожидаемое снижение эффекта тренировки. |
| | + | == Читайте также == |
| | | | |
| − | Гормоны щитовидной железы, или тироидные гормоны, оказывают сильное регуляторное воздействие на соотношение различных изоформ тяжелой цепи миозина в составе скелетной мышцы (D’Albis, Butler-Browne, 1993). Показано, что регуляция соотношения МуНС в скелетных мышцах крысы является специфической для пола и типа мышц (Larsson, Yu, 1997). Применение 3.5.3’-трийодтиронина (Т3) приводит к подавлению содержания MyHCI и увеличению содержания MyHCIIA в камбаловидной мышце самцов и самок, тогда как стимуляция содержания МуНСИХ наблюдалась только в мышцах самцов крыс (Larsson, Yu, 1997). Применение Т3 не вызывало никаких изменений в длинном разгибателе пальцев стопы у самцов крыс. В то же время в аналогичной ситуации в той же мышце у самок отмечалось достоверное изменение соотношения изоформ MyHCIIA и ИВ в пользу увеличения последней (Larsson, Yu, 1997). В целом эти результаты показывают, что сократительные качества скелетных мышц находятся под контролем ряда гормонов и ростовых факторов и изменение гормонального фона в этих мышцах при выполнении физических упражнений может быть в определенной степени ответственным за изменение характеристик мышцы в соответствии с физиологическими потребностями. Становится все более очевидным, что изменения структуры и функции мышц, происходящие под влиянием изменений гормонального фона, могут зависеть от пола и типа мышц.
| + | *[[Адаптация мышц к нагрузке]] |
| − | == Заключение ==
| + | *[[Адаптация мышц к тренировки на выносливость]] |
| − | | + | *[[Оценка адаптации организма на физические упражнения и тренировки]] |
| − | Были рассмотрены лишь отдельные аспекты значения специфических гормонов и ростовых факторов в регуляции некоторых важных параметров мышц, определяющих спортивные показатели. Эта сфера мышечной физиологии только начинает развиваться и здесь еще многое предстоит открыть прежде чем станет понятно взаимоотношение различных факторов, принимающих участие в разнообразных процессах адаптации скелетных мышц к различным видам двигательной активности. Последовательное описание различных этапов адаптации мышц к двигательной активности позволит создать основу для концепции индивидуализированного выбора упражнений с целью оптимизации качества тренировочных программ как для хорошо физически подготовленных лиц, так и для тех, кто ведет малоподвижный образ жизни, а также для специальных групп населения.
| + | *[[Адаптация к стрессорным ситуациям и физическим нагрузкам Меерсон Ф З]] |
| − | | + | [[Категория:Здоровье]] |
| − | == Литература ==
| |
| − | | |
| − | *Adams, G.R. & Haddad, F. (1996) The relationships among IGF-1, DNA content, and protein accumulation during skeletal muscle hypertrophy. Journal of Applied Physiology 81, 2509-2516. | |
| − | *Adams, G.R., Hather, B.M., Baldwin, K.M. & Dudley, G.A. (1993) Skeletal muscle myosin heavy chain composition and resistance training. Journal of Applied Physiology 74, 911-915.
| |
| − | *Allen, D.L., Monke, S.R., Talmadge, R.J., Roy, R.R. & Edgerton, V.R. (1995) Plasticity of myonuclear number in hypertrophied and atrophied mammalian skeletal muscle fibers. Journal of Applied Physiology 78, 1969-1976.
| |
| − | *Allen, D.L., Yasui, W., Tanaka, T. et al. (1996) Myonuclear number and myosin heavy chain expression in rat soleus single muscle fibers after spaceflight. Journal of Applied Physiology 81, 145-151.
| |
| − | *Allen, D.L., Roy, R.R. & Edgerton, V.R. (1999) Myonuclear domains in muscle adaptation and disease. Muscle and Nerve 22, 1350-1360. Andersen, J.L. & Aagaard, P. (2000) Myosin heavy chain IPX overshoot in human skeletal muscle. Muscle and Nerve 23, 1095-1104. Andersen, J.L., Klitgaard, H. & Sal tin, B. (1994) Myosin heavy chain isoforms in single fibres from m. vastus lateralis of sprinters: influence of training. Acta Physiologica Scandinavica 151, 135-142. Andersen, P. & Henriksson, J. (1977) Capillary supply of the quadriceps femoris muscle of man: adaptive response to exercise. Journal of Physiology 270, 677-690.
| |
| − | *Antonio, J. & Gonyea, WJ. (1993) Skeletal muscle fiber hyperplasia. Medicine and Science in Sports and Exercise 25, 1333-1345. Aroniadou-Anderjaska, V., Lemon, P.W., Gilloteaux, J. (1996) Effects of exogenous growth hormone on skeletal muscle of young female rats. Tissue and Cell 28, 719-724.
| |
| − | *Baker, J., Liu, J.P., Robertson, E.J. & Efstratiadis, A. (1993) Role of insulin-like growth factors in embryonic and postnatal growth. Cell 75, 73-82.
| |
| − | *Bamman, M.M., Shipp, J.R., Jiang, J. et al. (2001) Mechanical load increases muscle IGF-I and androgen receptor mRNA concentrations in humans. American Journal of Physiology 280, 383-390.
| |
| − | *Baumann, H., Jaggi, М., Sol and, F., Howald, H. & Schaub, M.C. (1987) Exercise training induces transitions of myosin isoform subunits within histochemically typed human muscle fibres. Pfingers Arcfiiv 409, 349-360.
| |
| − | *Bhasin, S., Storer, T.W., Berman, N. et al. (1996) The effects of sup-raphysiologic doses of testosterone on muscle size and strength in normal men. New England Journal of Medicine 335, 1-7.
| |
| − | *Biolo, G.f Fleming, R.Y., Maggi, S.P. & Wolfe, R.R. (1995) Transmembrane transport and intracellular kinetics of amino acids in human skeletal muscle. American Journal of Physiology 268, E75-E84.
| |
| − | *Biolo, G., Tipton, K.D., Klein, S. & Wolfe, R.R. (1997) An abundant supply of amino acids enhances the metabolic effect of exercise on muscle protein. American Journal of Physiology 273, E122-E129. Bischoff, R., 1994. The satellite cell and muscle regeneration. In: Myology (Engel, A.G., Franzini-Armstrong, C, eds.). McGraw-Hill, New York: 97-117.
| |
| − | *Boissonneault, G., Gagnon, J., Ho-Kim, M.A. & Tremblay, R.R. (1987) Lack of effect of anabolic steroids on specific mRNAs of skeletal muscle undergoing compensatory hypertrophy. Molecular and Cellular Endocrinology 51, 19-24.
| |
| − | *Bottinelli, R., Betto, R., Schiaffino, S. & Reggiani, C. (1994a) Maximum shortening velocity and coexistence of myosin heavy chain isoforms in single skinned fast fibres of rat skeletal muscle. Journal of Muscle Research and Cell Motility 15, 413-419.
| |
| − | *Bottinelli, R., Betto, R., Schiaffino, S. & Reggiani, C. (1994b) Unloaded shortening velocity and myosin heavy chain and alkali light chain isoform composition in rat skeletal muscle fibres. Journal of Physiology 478, 341-349.
| |
| − | *Brahm, H., Piehl-Aulin, K., Saltin, B. & Ljunghall, S. (1997) Net fluxes over working thigh of hormones, growth factors and biomarkers of bone metabolism during short lasting dynamic exercise. Calcified Tissue International 60, 175-180.
| |
| − | *Bricout, V., Germain, P., Serrurier, B. & Guezennec, C. (1994) Changes in testosterone muscle receptors: effects of an androgen treatment on physically trained rats. Cellular and Molecular Biology 40, 291-294.
| |
| − | *Cabric, M. & James, N.T. (1983) Morphometric analyses on the muscles of exercise trained and untrained dogs. American Journal of Anatomy 166, 359-368.
| |
| − | *Campion, D.R. (1984) The muscle satellite a review. International Review of Cytology 87 , 225-251.
| |
| − | *Charifi, N.. Kadi, F., Feasson, L. & Denis, C. (2003a) Effects of endurance training on satellite cell frequency in skeletal muscle of old men. Muscle and Nerve 28, 87-92.
| |
| − | *Charifi, N., Kadi, F., Feasson, L. et ah (2003b) Enhancement of microvessel tortuosity in the vastus lateralis muscle of old men in response to endurance training. Journal of Physiology 554, 559-569.
| |
| − | *Cheek, D.B. (1985) The control of cell mass and replication. The DNA unit: a personal 20-year study. Early Human Development 12, 211-239.
| |
| − | *Chesley, A., MacDougall J.D., Tarnopolsky, M.A., Atkinson, S.A. & Smith, K. (1992) Changes in human muscle protein synthesis after resistance exercise. Journal of Applied Physiology 73, 1383-1388.
| |
| − | *Clarke, M.S. & Feeback, D.L. (1996) Mechanical load induces sarcoplasmic wounding and FGF release in differentiated human skeletal muscle cultures. FASEB Journal 10, 502-509.
| |
| − | *Czesla, Mv Mehlhom, G., Fritzsche, D. & Asmussen, G. (1997) Cardiomyoplasty: improvement of muscle fibre type transformation by an anabolic steroid (metenolone). Journal of Molecular and Cellular Cardiology 29, 2989-2996.
| |
| − | *D’albis, A. & Butler-Browne, G. (1993) The hormonal control of myosin isoform expression in skeletal muscle of mammals: a review. Basic and Applied Myology 3(1), 7-16.
| |
| − | *Daugaard, J.R., Bramnert, М., Manhem, P. et al. (1999) Effect of 6 months of GH treatment on myosin heavy chain composition in GH-deficient patients. European Journal of Endocrinology 141, 342-349.
| |
| − | *DeChiara, T.M., Efstratiadis, A. & Robertson, EJ. (1990) A growth-deficiency phenotype in heterozygous mice carrying an insulin-like growth factor II gene disrupted by targeting. Nature 345, 78-80.
| |
| − | *Deschenes, M.R., Maresh, CM., Armstrong, L.E. et al. (1994) Endurance and resistance exercise induce muscle fiber type specific responses in androgen binding capacity. Journal of Steroid Biochemistry and Molecular Biology 50, 175-179.
| |
| − | *Dimauro, J., Balnave, R.J. & Shorey, CD. (1992) Effects of anabolic steroids and high intensity exercise on rat skeletal muscle fibres and capillarization. A morphometric study. European Journal of Applied Physiology 64, 204-212.
| |
| − | *Dionne, I,J., Kinaman, K.A. & Poehlman, E.T. (2000) Sarcopenia and muscle function during menopause and hormone-replacement therapy. Journal of Nutrition, Health and Aging 4(3), 156-161.
| |
| − | *Dodson, M.V., Allen, R.E. & Hossner, K.L. (1985) Ovine somatomedin, multiplication-stimulating activity, and insulin promote skeletal muscle satellite cell proliferation in vitro. Endocrinology 117. 2357-2363.
| |
| − | *Dorlochter, М., Astrow, S.H. & Herrera, A.A. (1994) Effects of testosterone on a sexually dimorphic frog muscle: repeated in vivo observations and androgen receptor distribution. Journal of Neurobiology 25, 897-916.
| |
| − | *Doumit, M.E., Cook, D.R. & Merkel, R.A. (1996) Testosterone up-regulates androgen receptors and decreases differentiation of porcine myogenic satellite cells in vitro. Endocrinology 137, 1385-1394.
| |
| − | *Egginton, S. (1987) Effects of an anabolic hormone on striated muscle growth and performance. Pflugers Archio 410, 349-355.
| |
| − | *Fernandez, A.M., Dupont, J., Farrar, R.P. et al. (2002) Muscle-specific inactivation of the IGF-I receptor induces compensatory hyperplasia in skeletal muscle. Journal of Clinical Investigation 109, 347-355.
| |
| − | *Florini, J.R., Ewton, D.Z. & Roof, S.L. (1991) Insulln-like growth factor-I stimulates terminal myogenic differentiation by induction of myogenin gene expression. Molecular Endocrinology (Baltimore, Md.) 5, 718-724.
| |
| − | *Floss, Т., Arnold, H.H. & Braun, T. (1997) A role for FGF-6 in skeletal muscle regeneration. Genes and Development 11, 2040-2051.
| |
| − | *Frederickson, R. & Sonenberg, N.. 1993. eIF-4E phosphorylation and the regulation of protein synthesis. In: Translational Regulation of Gene Expression (Ilan, J., ed.). Plenum Press, New York: 143-162.
| |
| − | *Fritzsche, D., Krakor, R., Asmussen, G. et al. (1994) Effect of an anabolic steroid (Metenolon) on contractile performance of the chronically stimulated latissimus dorsi in sheep. European Journal of Cardio-thoracic Surgery 8, 214-219.
| |
| − | *Fry, A.C, Allemeier, C.A. & Staron, R.S. (1994) Correlation between percentage fiber type area and myosin heavy chain content in human skeletal muscle. European Journal of Applied Physiology 68, 246-251.
| |
| − | *Galavazi, G. & Szirmai, J.A. (1971) Cytomorphometry of skeletal muscle: the influence of age and testosterone on the rat m. levator ani. Zeitschrift fur Zellforschung und Mikroskopische Anatomic 121, 507-530.
| |
| − | *Gerber, H.P., Condorelli, F., Park, J. & Ferrara, N. (1997) Differential transcriptional regulation of the two vascular endothelial growth factor receptor genes. Fit-1, but not Flk-1 / KDR, is up-regulated by hypoxia. Journal of Biological Chemistry 272, 23 659-23 667.
| |
| − | *Giddings, CJ. & Gonyea, W.J. (1992) Morphological observations supporting muscle fiber hyperplasia following weight-lifting exercise in cats. Anatomical Record 233, 178-195.
| |
| − | *Goldberg, A.L., Et linger, J.D., Goldspink, D.F. & Jablecki, С (1975) Mechanism of work-induced hypertrophy of skeletal muscle. Medicine and Science in Sports 7, 185-198.
| |
| − | *Goldspink, D.F., Cox, V.M., Smith, S.K. et al. (1995) Muscle growth in response to mechanical stimuli. American Journal of Physiology 268, 288-297.
| |
| − | *Goldspink, G. (1999) Changes in muscle mass and phenotype and the expression of autocrine and systemic growth factors by muscle in response to stretch and overload. Journal of Anatomy 194, 323-334.
| |
| − | *Goldspink, G., Scutt, A., Martindale, J. et al. (1991) Stretch and force generation induce rapid hypertrophy and myosin isoform gene switching In adult skeletal muscle. Biochemical Society Transactions 19, 368-373.
| |
| − | *Greeves, J.P., Cable, N.T., Reilly, T. & Kingsland, C. (1999) Changes in muscle strength in women following the menopause: a longitudinal assessment of the efficacy of hormone replacement therapy. Clinical Science (London, England) 97(1), 79-84.
| |
| − | *Gustafsson, Т., Puntschart, A., Kaijser, L., Jansson, E. & Sundberg, CJ. (1999) Exercise-induced expression of angiogenesis-related transcription and growth factors in human skeletal muscle. American Journal of Physiology 276, 679-685.
| |
| − | *Gustafsson, Т., Knutsson, A., Puntschart, A. et al. (2002) Increased expression of vascular endothelial growth factor in human skeletal muscle in response to short-term onelegged exercise training. Pflugers Archiv 444, 752-759.
| |
| − | *Hail, Z.W. & Ralston, E. (1989) Nuclear domains in muscle cells. Cell 59, 771-772.
| |
| − | *Hameed, М., Harridge, S.D. & Goldspink, G. (2002) Sarcopenia and hypertrophy: a role for insulin-like growth factor-I in aged muscle? Exercise and Sport Sciences Reviews 30, 15-19.
| |
| − | *Hameed, М., Orrell, R.W., Cobbold, М., Goldspink, G. & Harridge, S.D. (2003) Expression of IGF-I splice variants in young and old human skeletal muscle after high resistance exercise. Journal of Physiology 547 , 247-254.
| |
| − | *Hellsten, Y., Hansson, H.A., Johnson, L., Frandsen, U. & Sjodin, B. (1996) Increased expression of xanthine oxidase and insulin-like growth factor I (IGF-I) immunoreactivity in skeletal muscle after strenuous exercise in humans. Acta Physiologica Scandinavica 157, 191-197.
| |
| − | *Hickson, R.C. & Kurowski, T.G. (1986) Anabolic steroids and training. Clinics in Sports Medicine 5, 461-469.
| |
| − | *Hikida, R.S., Van-Nostran, S., Murray, J.D. et al. (1997) Myonuclear loss in atrophied soleus muscle fibers. Anatomical Record 247, 350-354.
| |
| − | *Hikida, R.S., Walsh, S., Barylski, N. et al. (1998) Is hypertrophy limited in elderly muscle fibers? A comparison of elderly and young strength-trained men. Basic and Applied Myology 8(6), 419-427.
| |
| − | *Hudlicka, O., Brown, M. & Egginton, S. (1992) Angiogenesis in skeletal and cardiac muscle. Physiological Reviews 72, 369-417.
| |
| − | *Inoue, K., Yamasaki, S., Fushiki, T. et al. (1993) Rapid increase in the number of androgen receptors following electrical stimulation of the rat muscle. European Journal of Applied Physiology 66, 134-140.
| |
| − | *Inoue, K., Yamasaki, S., Fushiki, Т., Okada, Y. & Sugimoto, E. (1994) Androgen receptor antagonist suppresses exercise-induced hypertrophy of skeletal muscle. European Journal of Applied Physiology 69, 88-91.
| |
| − | *Janssen, P.J., Brinkmann, A.O., Boersma, W.J. & Van-Der-Kwast, Т.Н. (1994) Immunohistochemical detection of the androgen receptor with monoclonal antibody F39.4 in routinely processed, paraffin-embedded human tissues after microwave pre-treatment. Journal of Histochemistry and Cytochemistry 42, 1169-1175.
| |
| − | *Jennische, E. (1989) Sequential immunohistochemical expression of IGF-I and the transferrin receptor in regenerating rat muscle in vivo. Acta Endocrinologica 121, 733-738.
| |
| − | *Jennische, E. & Matejka, G.L. (1992) IGF-I binding and IGF-I expression in regenerating muscle of normal and hypophysectomized rats. Acta Physiologica Scandinavica 146, 79-86.
| |
| − | *Jennische, E., Skottner, A. & Hansson, H.A. (1987) Satellite cells • express the trophic factor IGF-I in regenerating skeletal muscle. Acta Physiologica Scandinavica 129, 9-15.
| |
| − | *Joubert, Y. & Tobin, C. (1989) Satellite cell proliferation and increase in the number of myonuclei induced by testosterone in the levator ani muscle of the adult female rat. Developmental Biology 131, 550-557.
| |
| − | *Joubert, Y. & Tobin, C. (1995) Testosterone treatment results in quiescent satellite cells being activated and recruited into cell cycle in rat levator ani muscle. Developmental Biology 169, 286-294.
| |
| − | *Kadi, F. (2000) Adaptation of human skeletal muscle to training and anabolic steroids. Acta Physiologica Scandinavica 646, 1-52.
| |
| − | *Kadi, F. & Thomell, L.E. (1999) Training affects myosin heavy chain phenotype in the trapezius muscle of women. Histochemistry and Cell Biology 112, 73-78.
| |
| − | *Kadi, F. & Thomell, L.E. (2000) Concomitant increases in myonuclear and satellite cell content in female trapezius muscle following strength training. Histochemistry and Cell Biology 113, 99-103.
| |
| − | *Kadi, F., Eriksson, A., Holmner, S., Butler-Browne, G.S. & Thomell, L.E. (1999a) Cellular adaptation of the trapezius muscle in strength-trained athletes. Histochemistry and Cell Biology 111, 189-195.
| |
| − | *Kadi, F., Eriksson, A., Holmner, S. & Thomell, L.-E. (1999b) Effects of anabolic steroids on the muscle cells of strength-trained athletes. Medicine and Science in Sports and Exercise 31, 1528-1534.
| |
| − | *Kadi, F., Ahlgren, C, Waling, K., Sundelin, G. & Thomell, L.E. (2000a) The effects of different training programs on the trapezius muscle of women with work-related neck and shoulder myalgia. Acta Neuropathologica 100, 253-258.
| |
Источник:
Учебное пособие для ВУЗов «Спортивная физиология».
Автор: И.И. Земцова Изд.: Олимпийская лит-ра, 2010 год.
Адаптация как биологическая основа эффектов занятий физическими упражнениями[править | править код]
Адаптация — это способность организма приспосабливаться к внешней среде или изменениям в самом организме. Различают процесс и состояние адаптации. Состояние адаптации — это состоявшаяся физиологическая адаптация. Оно характеризуется устойчивым уровнем активности и взаимосвязи систем органов, тканей и механизмов регуляции, обеспечивающих нормальный уровень жизнедеятельности организма в новых условиях внешней и внутренней среды. Это состояние достигается в течение определенного времени, за которое происходит адаптация (процесс адаптации) (Ключевые факторы адаптации..., 1986; Медведев, 1984; Солодков, Сологуб, 2003).
По механизмам возникновения различают такие виды адаптации:
- генотипическая, обусловленная врожденными механизмами функционирования систем и возникшая на основе наследственности, изменчивости, естественного отбора, мутаций и являющаяся характерной для вида живых организмов;
- фенотипическая, приобретаемая в течение индивидуальной жизни и проявляемая относительно влияния определенного фактора внешней среды (температура, АД, влажность, физические нагрузки и др.).
По срокам возникновения различают срочную (быструю, незавершенную) и долговременную (медленную, завершенную) фазы адаптации.
Срочная адаптация — это реакция организма на действующий раздражитель. Ее реализация происходит на основе сформировавшихся ранее физиологических механизмов. Адаптация реализуется «с места» и не является совершенной. Примером срочной адаптации являются физиологические реакции организма на выполнение физической нагрузки: повышение ЧСС, ЧД, АД и других показателей. Работа требует затрат энергии, и для ее реализации усиливается деятельность вегетативных систем.
Долговременная адаптация возникает в результате многократного влияния фактора, то есть многократной реализации срочной адаптации. Вследствие постепенного количественного накопления изменений организм приобретает новое качество — превращается в адаптированный, и потому одни и те же движения выполняются экономнее, эффективнее, а максимальная работа выводит организм на более высокий уровень функционирования.
Выполнение оптимальных, адекватных индивидуальным возможностям организма, физических нагрузок обеспечивает развитие структурных изменений прогрессивного характера — рациональной адаптации, что способствует развитию резервных возможностей организма спортсменов. Чрезмерные физические нагрузки, превышающие функциональные возможности организма, вызывают нарушение функциональных связей между его структурными уровнями и ведут к развитию нерациональной адаптации. В отличие от рациональной, нерациональная адаптация характеризуется стремительным становлением и может сопровождаться дистрофическими изменениями тканей, а впоследствии и структурно-функциональными нарушениями органов и тканей (Алексаняйц, 2003; Козлов, 1997; Спортивная медицина..., 2003).
Срочная адаптация рассматривается как состояние общего напряжения организма, возникающее вследствие влияния очень сильного раздражителя. Термин «стресс» впервые использовал канадский ученый Ганс Селье в 1936 г. Он показал, что во время влияния на организм стрессового раздражителя возникает активизация деятельности организма, увеличивающая секрецию АКТГ, которая стимулирует прежде всего деятельность коры надпочечников.
Гормоны коры надпочечников стимулируют приспособительные механизмы, благодаря которым организм адаптируется к влиянию раздражителя. Механизмы такой срочной адаптации являются общими для разных стрессовых влияний — физических, химических, эмоциональных и др. В результате возникло понятие — общий адаптационный синдром (Дубровский, 2005; Фомин, Вавилов, 1991).
Общий адаптационный синдром — это комплекс неспецифических реакций организма на действие раздражителя, происходящих в несколько стадий: тревоги, резистентности, истощения.
Стадия тревоги характеризуется предельной мобилизацией функций. Физиологические механизмы этой стадии характерны для срочной адаптации, реализующейся по типу стресс-реакции с максимальной мобилизацией вегетативных функций.
Стадия резистентности характеризуется активным поиском устойчивого состояния. Ее механизмы являются основой долговременной адаптации.
Стадия истощения проявляется в случае, когда сила действующего раздражителя продолжает расти, превышая функциональные и метаболические возможности организма, возникает срыв адаптации, переходящий в дезадаптацию. При оптимальной организации тренировочного процесса стадия резистентности может не возникать в течение продолжительного времени.
Действие закона адаптации в процессе тренировки спортсменов (по Zatsiorsky, 1995)
Целенаправленный тренировочный процесс приводит к разнообразным изменениям в организме спортсмена и, таким образом, увеличивает их работоспособность. С биологической точки зрения тренировка - это длительный процесс адаптации спортсмена к различным нагрузкам. Следовательно, упражнения, тренировки и различные задания служат стимулами для адаптации. В биологии адаптация рассматривается как процесс приспособления, который происходит в организме под воздействием изменяющихся условий жизни. Вообще говоря, адаптация, первоначально описанная великим физиологом Гансом Селье (1950), является одним из фундаментальных законов науки о жизни. Профессор В.М. Зациорский применил принципы процесса адаптации к спортивной тренировке (1995); он обнаружил, что приспособление спортсменов к увеличивающимся нагрузкам обусловлено тремя главными факторами: величиной воздействия, его спецификой и приспособляемостью спортсмена (рис.).
По закону адаптации эффективная тренировка должна обеспечивать оптимальную комбинацию этих трёх главных факторов, а она, в свою очередь, определяет прогресс в работе над работоспособностью спортсменов. Обобщая сказанное, можно назвать упомянутые выше факторы принципами адаптации применительно к тренировочному процессу.
Величина тренировочной нагрузки и принцип перегрузки[править | править код]
Тренировочная нагрузка вызывает реакцию спортсмена и служит стимулом для адаптации. Величина воздействия может регулироваться тремя факторами: объёмом нагрузки, её интенсивностью и новизной упражнений. Важно отметить, что рост уровня подготовленности может быть достигнут, только если величина воздействия достаточна. Принцип перегрузки гласит, что для увеличения уровня подготовленности требуется применение нагрузки (воздействия), величина которой превышает привычный уровень.
В соответствии с принципом перегрузки величина нагрузки имеет первостепенное значение и должна тщательно оцениваться и программироваться. Общий подход к описанию величины нагрузки представлен ниже (табл.).
Характеристики величины нагрузки
|
Компонент тренировочной нагрузки |
Основные показатели |
Возможные индикаторы |
|
Объём |
Сумма всех выполненных упражнений, представленная количественной характеристикой |
Общее количество тренировок за период времени, например, за неделю, месяц, год и т.д.
Общее время, затраченное на тренировки за данный период.
Общий километраж за тренировочный период.
Общее количество подъёмов, бросков, прыжков и пр. за тренировочный период |
|
Интенсивность |
1) Интенсивность рабочей нагрузки.
2) Сумма упражнений, выполненных с увеличенной мощностью |
Уровень мощности (%) относительно максимума.
Уровень мощности, определяемый по величине ЧСС.
Соответствие определённой зоне интенсивности. Частный объём упражнений, выполненных с увеличенной мощностью (километраж, затраченное время, количество попыток и т.д.) |
|
Новизна
упражнения |
Наличие упражнения, которое содержит неизвестные элементы или детали / новые комбинации известных элементов |
Количество новых (или относительно новых) упражнений, включённых в программу тренировки |
Объём тренировочной нагрузки. Исторически самый простой способ увеличить нагрузку состоял в увеличении объёма тренировок. У высококвалифицированных спортсменов во многих видах спорта в 1930-х годах количество тренировок в неделю равнялось 2-3, в 1960-х увеличилось до 6-8, а в 1980-х достигло 9-14. С тех пор частота тренировок осталась на том же уровне. В течение долгого времени считалось, что желание увеличить объём тренировок было ограничено физиологическими и социальными факторами. С точки зрения физиологов уже был достигнут верхний предел резервов человека; социологи же высказывали беспокойство о том, что кроме тренировок спортсмены нуждаются в образовании, профессии, личной жизни и т.д.
Несмотря на это, объём тренировочных нагрузок в мировом спорте стремился к увеличению до конца 1980-х годов. Этот объём стабилизировался и даже уменьшился только в течение двух прошедших десятилетий. В любом случае, увеличение тренировочной нагрузки - слишком очевидный фактор личного прогресса спортсмена в любом виде спорта. Оценка объёма тренировочной нагрузки - обычная практика в видах спорта на выносливость, где выполненный километраж традиционно подсчитывается, однако это может стать трудной задачей в игровых видах или в единоборствах, где не просто суммировать количество специфических спортивных действий.
Интенсивность тренировочной нагрузки. Интенсивность тренировочной нагрузки обычно рассматривается в двух аспектах:
- как мерило уровня мощности относительно максимума (иногда относительно уровня соревновательной мощности);
- как составляющая общего объёма тренировочной нагрузки, который выполнен с увеличенной (выше обычного) мощностью.
Конечно, более интенсивные упражнения вызывают более явную реакцию в организме спортсмена. Следовательно, интенсивность нагрузки оценивается как показателями внешней нагрузки (скоростью, мощностью, поднятыми весами), так и посредством индикаторов реакции организма. Частота сердечных сокращений (ЧСС), например, является одним из широко распространённых показателей физиологической реакции. ЧСС обеспечивает достаточную индикацию уровня интенсивности широкого спектра упражнений.
В последние годы зоны интенсивности (ЗИ) стали широко использоваться во многих видах спорта и для планирования, и для посттренировочной оценки (Viru, 1995). В соответствии с этим подходом весь диапазон интенсивности подразделяется на зоны (обычно их пять). Каждая ЗИ описывается рядом значимых индикаторов, каждый из которых отражает диапазон показателей, соответствующих этой зоне. Обычно для характеристики определённой зоны интенсивности используются лактат крови, ЧСС, скорость (или время работы, или мощность) и темп движений. За прошедшее десятилетие в связи с развитием новых спортивных технологий (таких как мониторы ЧСС, портативные анализаторы лактата крови, электронные измерительные системы времени) этот подход был существенно усовершенствован.
Новизна упражнения. Новизна упражнения является третьим компонентом, определяющим величину тренировочной нагрузки; реакция спортсменов весьма зависима от того, насколько привычными являются для них некоторые упражнения. Однако в отличие от объёма и интенсивности новизна упражнения редко рассматривается как фактор, влияющий на тренировочную нагрузку. Известно, что творчески настроенные тренеры повсюду ищут новые оригинальные упражнения, чтобы обогатить существующий набор и сделать тренировочный процесс более привлекательным. Эффект применения этих новшеств проявляется в более выраженной физиологической реакции спортсмена.
Пример. Игорь Кошкин (СССР), один из всемирно известных экспертов в области плавания, который тренировал трёхкратного олимпийского чемпиона Владимира Сальникова, сказал другим тренерам: «Если Вы начнёте использовать стояние на голове как упражнение для ваших пловцов, начальный эффект будет существенным и положительным из-за его новизны. Но этот эффект будет очень кратковременным, потому что это упражнение не затрагивает специфические плавательные способности ваших спортсменов».
Это замечание подчёркивает сложность проблемы, связанной с новизной упражнения. Действительно, нетрудно найти упражнение, с которым спортсмены не знакомы, но нелегко найти незнакомое им упражнение, которое соответствует специфическим по виду спорта физиологическим, биомеханическим и психологическим требованиям. Именно поэтому специфичность тренировочной нагрузки, которая будет рассмотрена ниже, является обязательным фактором адаптации в спортивной тренировке.
Специфичность тренировочной нагрузки[править | править код]
Как видно из рисунка, специфичность тренировочной нагрузки характеризуется переносом результата тренировки с одного задания (выполнения вспомогательного упражнения) на другое (основное упражнение). Обычно тренеры используют широкий набор упражнений, большинство из которых может быть разделено на две группы:
- упражнения для совершенствования физических качеств (силы, выносливости и др.);
- упражнения для совершенствования технических навыков.
Конечно, эти упражнения могут быть скомбинированы, чтобы улучшить взаимодействие между физическими качествами и техническими навыками. В любом случае, полезность каждого упражнения зависит от того, как оно влияет на выполнение главного (соревновательного) упражнения. Другими словами, перенос двигательных способностей и перенос технических навыков с тренировочного упражнения на соревновательное определяет, насколько полезными являются эти вспомогательные упражнения.
Две тесно связанные особенности характеризуют аккомодацию (обязательный компонент процесса адаптации, вызванного тренировкой):
- увеличение работоспособности;
- уменьшение реакции на стандартную физическую нагрузку.
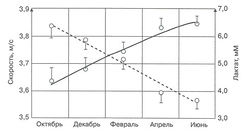
Изменение скорости анаэробного порога (сплошная линия) и накопление лактата крови после прохождения 1000-метровой дистанции с заранее определённой скоростью (пунктирная линия) у квалифицированных байдарочников в ходе подготовительного периода
Увеличение работоспособности может быть отслежено с помощью специфических по виду спорта показателей, таких как результаты выполнения работы до отказа, скорость анаэробного порога в видах спорта на выносливость и т.д. Стандартная физическая нагрузка может быть организована при обследовании спортсменов на эргометре или при тестировании их с заранее заданной скоростью или мощностью. Оба варианта можно проследить на примере результатов наблюдения за подготовкой байдарочников мирового класса в течение одного сезона.
Конкретный пример. Группа из девяти квалифицированных байдарочников обследовалась в течение одного подготовительного сезона. Выполнялся тест на воде (4 раза по 1000 м) с возрастающей нагрузкой для определения скорости анаэробного порога. Кроме того, выполнялась стандартная нагрузка - гребля на дистанции 1000 м на заранее определённой скорости, соответствующей средней интенсивности (скорость была запрограммирована лидирующей моторной лодкой). После испытания брались пробы крови. Графики показывают значительное увеличение скорости анаэробного порога у всех спортсменов и аналогичное уменьшение накопления лактата крови, вызванное греблей с постоянной скоростью (рис.).
Этот пример показывает, что процесс аккомодации у спортсменов может контролироваться посредством испытаний, выполняемых как с максимальными, так и со стандартными усилиями. Этот подход может также использоваться в видах спорта, где спортивный результат не поддаётся измерению, таких как игры с мячом, где стандартная нагрузка может быть запрограммирована определённой комбинацией специфических по виду спорта элементов с фиксированной частотой выполнения и диапазоном движения.
У процесса аккомодации есть много субъективных индикаторов: с увеличением работоспособности спортсмены сообщают о большей «свободе движения», облегчении дыхания во время длительной работы, лучшем расслаблении мышц, усилении специфических по виду спорта ощущений, подобных «чувству воды» в водных видах, «чувству льда» в катании на коньках и т.д. Все эти субъективные оценки очень важны и для тренера, и для спортсмена; желательно отмечать их в дневниках спортсменов и журналах тренеров.
В заключение надо отметить, что общая логика принципов адаптации может быть представлена в такой последовательности:
- тренировка с адекватной рабочей нагрузкой вызывает желаемые реакции в организме спортсменов (принцип величины воздействия),
- эти реакции вызывают процесс приспособления, который приводит к увеличению работоспособности и более экономному реагированию на стандартные рабочие нагрузки (принцип аккомодации);
- увеличенный уровень работоспособности отражается на выполнении соревновательного упражнения в соответствии с тренировочными результатами, перенесёнными с различных упражнений на главное соревновательное (принцип специфичности).
Нарушая эти взаимосвязи, мы понижаем тренировочный эффект, и чем выше уровень спортсмена, тем значительнее будет ожидаемое снижение эффекта тренировки.


